(132h) Engineering the Retinal Microenvironment to Improve Retinal Ganglion Cell Transplantation
AIChE Annual Meeting
2024
2024 AIChE Annual Meeting
Food, Pharmaceutical & Bioengineering Division
Faculty Candidate Session: Food, Pharmaceuticals, and Bioengineering I
Monday, October 28, 2024 - 2:36pm to 2:54pm
Glaucoma is the leading cause of irreversible blindness worldwide, disproportionally affecting those of lower socioeconomic status and people over the age of 40.1 Glaucoma and other optic neuropathies are characterized by the loss of retinal ganglion cells (RGCs), the neurons in the retina that relay all visual information from the eye to the brain. While available treatments can slow disease progression,2 there are currently no therapies to restore lost visual function and RGCs1. The mammalian retina does not possess the capacity to regenerate; thus, RGC death leads to irreversible vision loss.3
Cell transplantation is a promising solution to replace RGCs lost in advanced retinal disease.4 However, poor survival rates and donor cell integration have limited the effectiveness and further development of RGC replacement therapies.4–7 One of the significant barriers to successful RGC integration into the existing neural circuitry is cell migration toward their natural position in the retina, the ganglion cell layer (GCL).4,5 Another is that the glaucomatous retina is typically characterized by increased inflammation, immune cell infiltration, and host cell reactivity,2,4,8 making it challenging for transplanted donor cells to survive in the host retina. To overcome these limitations and support donor neuron survival and integration, we establish a methodology for controlling stem cell-derived neurons in vivo by engineering their microenvironment post-transplantation.
Methods:
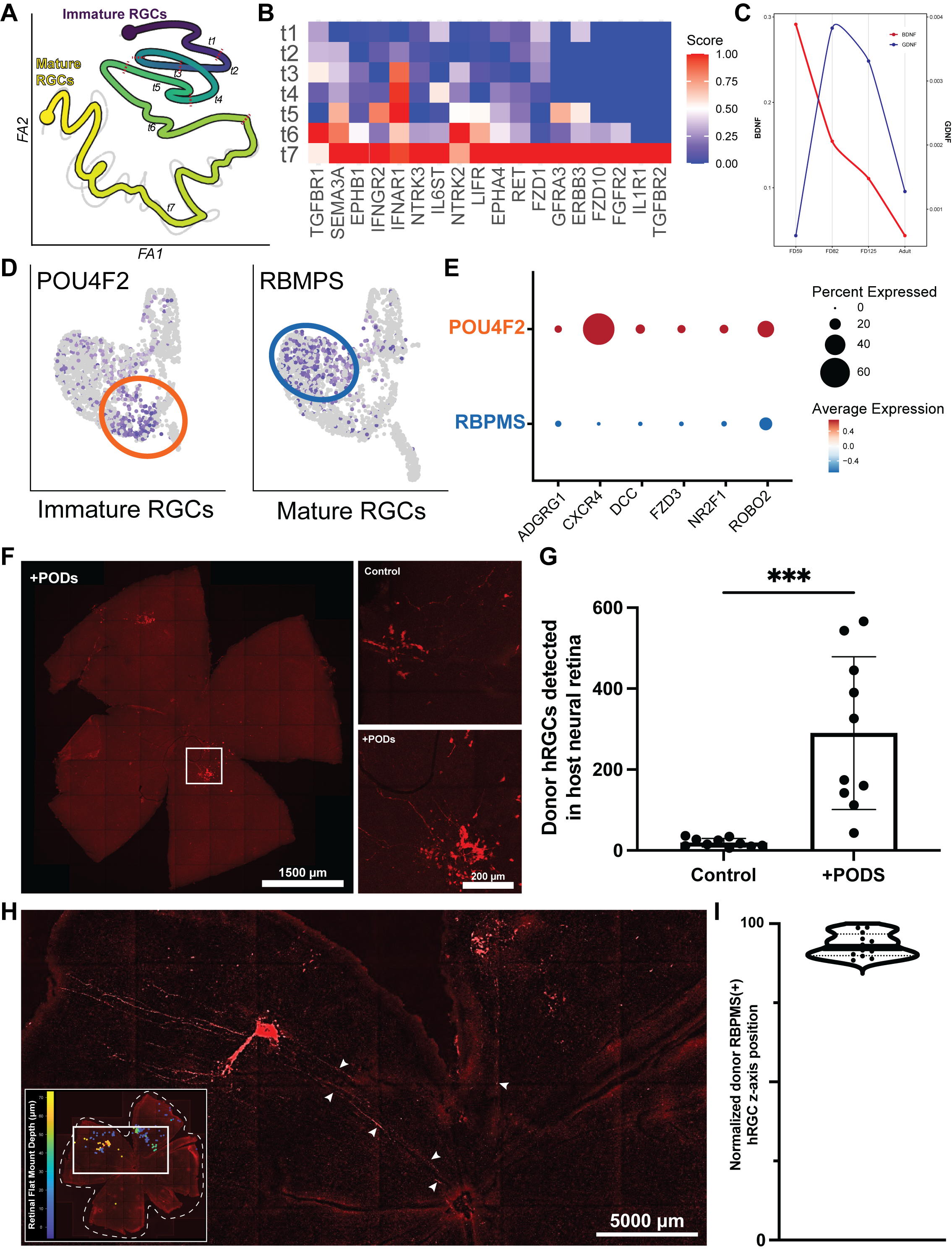
To identify factors that can be used to modify the retinal microenvironment and control donor neuron behavior, we first analyzed the single-cell transcriptome of the developing human retina in silico. Our in silico analysis identified six receptor-ligand candidates and 18 neurotropic factors that play a role in newborn neuron survival and migration during development (Fig. 1A-E). We hypothesized that a subset of these proteins may promote donor stem cell-derived RGC survival and migration in the retina.
We screen these candidates using functional assays (live/dead staining and a transwell migration assay) before testing the lead candidates in the transplantation setting to limit the number of animals. For this and other experiments, RGCs were differentiated from Brn3b-tdTomato human ESCs in 3D retinal organoid cultures.9–11
To support donor RGCs and establish a proneuronal microenvironment, we formulated human donor RGCs with slow-release neurotropic factors (GDNF- and BDNF-loaded polyhedrin-based particles (PODs), the most potent neurotropic factors for RGC survival).12 Donor RGCs were transplanted (20,000 RGCs/retina) into an RGC-deficient mouse retina (inherited loss or neurotoxic model) with and without PODs. To improve donor neuron migration in vivo, we transplanted RGCs subretinally and delivered recombinant SDF1 protein (the most potent chemokine for RGC recruitment in vitro) intravitreally to establish a chemokine gradient across the retina.11 Three days after transplantation, retinas were stained for host and donor cells to assess RGC integration and survival by tracking the position of each donor RGC in 3D reconstructions of retinal flat mounts (8 – 11 mice/group). Donor RGC maturation was assessed by RBPMS staining using these same retinas.
Results:
Our in silico analysis identified BDNF and GDNF as neurotrophic factors during development (Fig. 1A-C). These factors are essential during the early stages of RGC development, but they are not present in adulthood (Fig. 1C), indicating we may need to supplement the retina with these factors to establish an environment to support regeneration. Our transplantation experiments demonstrated that delivering BDNF/GDNF-PODS with donor RGCs significantly improved the transplantation outcome. Specifically, the PODs formulation led to a 15-fold increase in the total number of human donor RGCs detected in the mouse retina (Fig. 1F-G).
Furthermore, after identifying SDF1 as the most potent chemokine using our transwell migration assay, we demonstrate that treatment with SDF1 significantly increased donor RGC integration into the GCL from 16 ± 11% to 44 ± 11%. We confirmed the direct effect of SDF1 on donor cells by inhibiting in response to SDF1 by blocking SDF1-CXCR4 binding in donor cells with AMD3100, with only 17 ± 5.9% of donor RGC migrating into the GCL.
Lastly, we demonstrate that only donor RGCs that migrate into the GCL because of SDF1 treatment extend their axons towards the optic nerve head and express mature RGC markers (Fig. 1H-I). Further, the number of transplants showing donor RGC neurite extension toward the optic nerve head (average length of 1008.44 ± 264.12 µm) is significantly increased from 10% to 50% with PODs co-treatment. This finding is critical as it demonstrates that the transplanted cells have the potential to connect with their postsynaptic targets in the brain, a necessary step for functional recovery after injury.
Conclusion:
The combination of modulating the host retinal microenvironment with slow-release growth factor and chemokines is an effective tool for improving transplantation outcomes by improving donor RGC survival and structural integration. Specifically, BDNF and GDNF PODs improve donor RGC survival 15-fold, while an exogenous SDF1 gradient improves the structural integration of human stem cell-derived neurons 2.7-fold. While further research is necessary to improve donor RGC functional integration, modifying the retinal microenvironment may offer a viable solution to the challenge of RGC replacement.
In this study, we demonstrated that our “in silico—in vitro—in vivo†funnel-like approach can identify factors that can be used to engineer the tissue microenvironment to control donor RGC survival and migration. In the future, this approach can potentially be applied to control and improve any number of aspects of donor and newborn neuron behavior within the nervous systems. Altogether, our findings have significant implications for restoring lost visual function in glaucoma and other retinal diseases and highlight the potential of microenvironment engineering as a promising approach for future research and clinical applications.
References:
1. Tham, Y.-C. et al. Global Prevalence of Glaucoma and Projections of Glaucoma Burden through 2040 A Systematic Review and Meta-Analysis. Ophthalmology 121, 2081–2090 (2014).
2. Storgaard, L., Tran, T. L., Freiberg, J. C., Hauser, A. S. & Kolko, M. Glaucoma Clinical Research: Trends in Treatment Strategies and Drug Development. Frontiers Medicine 8, 733080 (2021).
3. Erskine, L. & Herrera, E. Connecting the Retina to the Brain. Asn Neuro 6, 1759091414562107 (2014).
4. Soucy, J. R. et al. Retinal ganglion cell repopulation for vision restoration in optic neuropathy: a roadmap from the RReSTORe Consortium. Mol. Neurodegener. 18, 64 (2023).
5. Zhang, K. Y., Aguzzi, E. A. & Johnson, T. V. Retinal Ganglion Cell Transplantation: Approaches for Overcoming Challenges to Functional Integration. Cells 10, 1426 (2021).
6. Venugopalan, P. et al. Transplanted neurons integrate into adult retinas and respond to light. Nat Commun 7, 10472 (2016).
7. Oswald, J., Kegeles, E., Minelli, T., Volchkov, P. & Baranov, P. Transplantation of miPSC/mESC-derived retinal ganglion cells into healthy and glaucomatous retinas. Mol Ther - Methods Clin Dev 21, 180–198 (2021).
8. Tehrani, S. et al. Astrocyte Structural and Molecular Response to Elevated Intraocular Pressure Occurs Rapidly and Precedes Axonal Tubulin Rearrangement within the Optic Nerve Head in a Rat Model. Plos One 11, e0167364 (2016).
9. Wahlin, K. J. et al. CRISPR Generated SIX6 and POU4F2 Reporters Allow Identification of Brain and Optic Transcriptional Differences in Human PSC-Derived Organoids. Frontiers Cell Dev Biology 9, 764725 (2021).
10. Sluch, V. M. et al. Enhanced Stem Cell Differentiation and Immunopurification of Genome Engineered Human Retinal Ganglion Cells. Stem Cell Transl Med 6, 1972–1986 (2017).
11. Soucy, J. R. et al. Controlling donor and newborn neuron migration and maturation in the eye through microenvironment engineering. Proc. Natl. Acad. Sci. 120, e2302089120 (2023).
12. Soucy, J. R. et al. Sustained Neurotrophic Factor Cotreatment Enhances Donor and Host Retinal Ganglion Cell Survival in Mice. BioRxiv 2024.03.07.583961 (2024) doi:10.1101/2024.03.07.583961.
Figure 1. Developmental relevant pro-survival and pro-migratory factors improve donor RGC transplantation. (A) Cell fate trajectory based on pseudo-time analysis of developing RGCs from immature (dark blue) to mature (yellow). (B) Average expression levels of all the receptors for the neurotrophic factor identified as possibility critical for cell survival during development. (C) BDNF and GDNF expression levels in the retina during different stages of development. (D) Feature plots showing characteristic genes describing clusters of mature (RBPMS) and immature (Pou4f2) RGCs in the developing retina. (E) Dot plot showing the expression of select receptors involved in neural migration identified as possibility critical for RGC migration during development. (F) Representative high magnification images from retinal whole mounts showing human donor RGCs (red) and (G) the number of donor RGCs in the host retina. (H) Representative retinal flat mount (max intensity projection) shows donor RGCs extending neurites toward the optic nerve head (white arrows) following subretinal cell delivery and intravitreal SDF1 treatment. The inlay shows a depth-coded retinal flat mount for all identified donor RGCs – those in the GCL are shown in yellow. (I) Single-cell quantification of the z-axis position normalized to the thickness of the retina reveals that only donor RGC that migrate into the GCL express RBPMS. Adapted from Soucy et al., PNAS, 2023 and Soucy et al., BioRxiv, 2024.11,12