(23e) Microfluidic Assembly of Mitochondria-Loaded Microparticles for on-Demand Delivery
AIChE Annual Meeting
2024
2024 AIChE Annual Meeting
Food, Pharmaceutical & Bioengineering Division
Cell and Tissue Engineering: Engineered Therapeutics
Sunday, October 27, 2024 - 5:04pm to 5:22pm
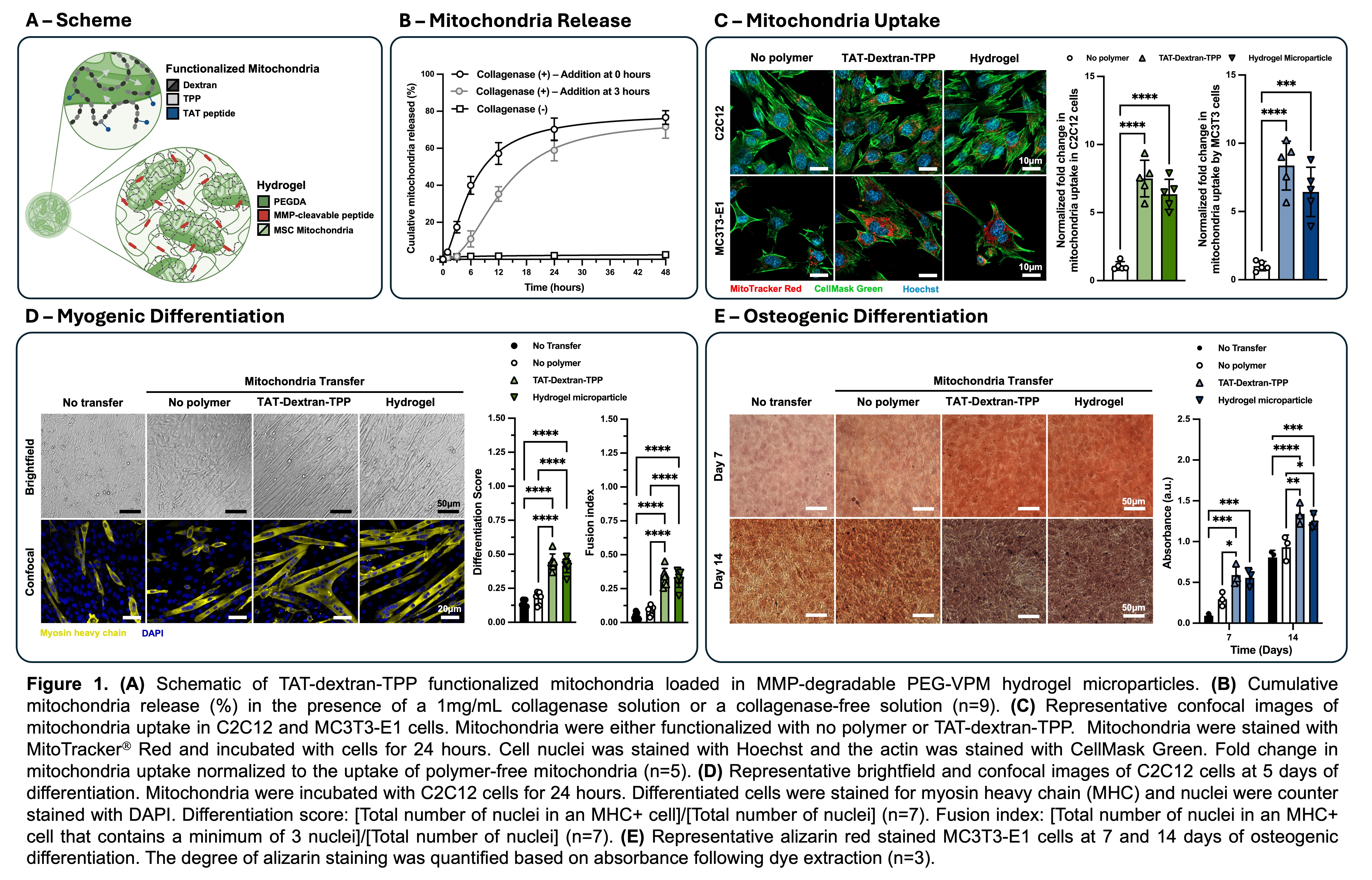
Methods. TAT-peptides (QPRRRQRRKKKRG), known to facilitate endocytosis of extracellular vesicles/particles, and triphenol phosphate (TPP), designed to self-insert into the mitochondrial membrane, were conjugated to dextran (TAT-dextran-TPP) and later used to surface functionalize mitochondria. The conjugation of TAT-peptide and TPP was calculated to be 10% and 40% respectively. Isolated mitochondria (stained with MitoTrackerÒ Red) were incubated with 2.5wt% TAT-dextran-TPP for 12 hours and subsequently encapsulated in MMP-cleavable hydrogels (20µg/mL total mitochondria protein) using droplet microfluidics. The hydrogel microparticles consisted of 10wt% poly(ethylene glycol) conjugated with the MMP-cleavable peptide GGVPMSMRGGGK (PEG-VPM). Enzyme-directed release of mitochondria was characterized in the presence of 1mg/mL collagenase. In parallel, either polymer-free mitochondria, TAT-dextran-TPP functionalized mitochondria, or TAT-dextran-TPP functionalized mitochondria loaded into PEG-VPM hydrogel microparticles were incubated with mouse myoblasts (C2C12 cells) or osteoprogenitor cells (MC3T3-E1 cells) for 24 hours. Following 24-hour incubation, mitochondria uptake was characterized using confocal microscopy processed with ImageJ. Following mitochondria transfer, the degree of myogenesis in C2C12 cells was visualized with myosin heavy chain (MHC) immunostaining at day 5 and quantified with a differentiation score and fusion index. Following mitochondria transfer, the degree of osteogenesis in MC3T3-E1 cells was visualized with alizarin red staining (ARS) at days 7 and 14 and quantified with a colorimetric assay.
Results & Discussion. MSC-derived mitochondria encapsulated in ~90µm diameter MMP-cleavable hydrogel microparticles were actively released over 48 hours when in the presence of collagenase (Figure 1B). Unlike small molecule drugs and proteins, mitochondria range from 500nm to 1µm in diameter, and therefore cannot actively diffuse through the pores of the gel. As such, no release was observed in a collagenase-free solution, illustrating selective release in the presence of MMPs.
In order to facilitate the uptake of exogenous mitochondria post release, the surface of the mitochondria were modified with TAT-dextran-TPP polymers. Transactivator of transcription (TAT) peptides are a class of cell penetrating peptides that actively promote endocytosis of conjugated cargos5. The uptake of MSC-derived mitochondria (stained with MitoTracker® Red) was quantified as the fold change in the average number of mitochondria positive pixels per cell relative to the polymer-free mitochondria transfer condition (Figure 1C). Cells were segmented using the actin stain CellMaskÔ Green and nuclei were counterstained with Hoechst 33342. We found a 7.5-fold and 6.3-fold increase in C2C12 uptake of TAT-dextran-TPP functionalized mitochondria and TAT-dextran-TPP functionalized mitochondria released from hydrogel microparticles respectively following 24-hour incubation. There was no statistical significance in uptake between the TAT-dextran-TPP functionalized mitochondria and TAT-dextran-TPP functionalized mitochondria released from hydrogel microparticles, indicating that the process to encapsulate and release mitochondria does not hinder the uptake by recipient cells. Similarly, we found an 8.4-fold and 6.4-fold increase in MC3T3-E1 uptake of TAT-dextran-TPP functionalized mitochondria and TAT-dextran-TPP functionalized mitochondria released from hydrogel microparticles respectively following 24-hour incubation. Again, there was no statistical significance in uptake between the TAT-dextran-TPP functionalized mitochondria and TAT-dextran-TPP functionalized mitochondria released from hydrogel microparticles.
While the increased uptake of mitochondria was exciting, we looked to prove that the degree of uptake was sufficient to enhance both myogenesis in C2C12 cells and osteogenesis in MC3T3-E1 cells. Following mitochondria transfer, C2C12 cells were cultured in FBS-depleted media for 5 days to facilitate myogenic differentiation. The cells were then immunostained for myosin heavy chain (MHC) as a marker for differentiation and nuclei were counterstained with DAPI (Figure 1D). Two different metrics were used to determine the degree of differentiation: Differentiation score ([total number of nuclei in an MHC+ cell]/[total number of nuclei]) and fusion index ([total number of nuclei in an MHC+ cell that contains a minimum of 3 nuclei]/[total number of nuclei]). After 5 days, we observed greater differentiation and greater fusion into myotubes for conditions with increased mitochondria uptake (TAT-dextran-TPP functionalized mitochondria and TAT-dextran-TPP functionalized mitochondria released from hydrogel microparticles). We also characterized the osteogenic differentiation of MC3T3-E1 cells post mitochondria transfer. Osteogenic differentiation of osteoprogenitor cells was performed over 14 days in 5 mM b-glycerophosphate and 50 mg/ml ascorbic acid supplemented a-MEM media. At days 7 and 14, an alizarin red assay was used to stain for calcium deposition and quantified through acetic acid extraction and neutralization with ammonium hydroxide followed by colorimetric detection at 405nm (Figure 1E). At days 7 and 14 we observed greater calcium deposition for conditions with increased mitochondria uptake (TAT-dextran-TPP functionalized mitochondria and TAT-dextran-TPP functionalized mitochondria released from hydrogel microparticles). In both the myogenic and osteogenic assays there was no statistical difference between the TAT-dextran-TPP functionalized mitochondria and TAT-dextran-TPP functionalized mitochondria released from hydrogel microparticles groups leading to the conclusion that release from the gels does not affect mitochondrial viability.
Conclusions. From this study, we are able to illustrate that mitochondria can be isolated from MSC culture, loaded into MMP-cleavable hydrogel microparticles and released, all while preserving their function in vitro. Moreover, mitochondria surface modification with TAT-dextran-TPP increases internalization by C2C12 and MC3T3-E1 cells, promoting a larger degree of myogenic and osteogenic differentiation respectively. We note that while these results are most directly applicable for muscle and bone regeneration, on-demand mitochondrial release for localized cellular uptake has applications in a wide range of regenerative medicine applications.
Acknowledgement. This work was supported by the Carol Ann and David D. Flanagan Professorship in BME at GT/Emory.
References.
[1] J.L. Spees, S.D. Olson, M.J. Whitney, and D.J. Prockop. Mitochondrial transfer between cells can rescue aerobic respiration. Proceedings of the National Academy of Sciences 2006, 103 (5), 1283-1288
[2] N. Borcherding and J.R. Brestoff. The power and potential of mitochondria transfer. Nature 2023, 623, 283-291
[3] J. Sun, H.T.J. Lo, L. Fan, T.L. Yiu, A. Shakoor, G. Li, W.Y.W. Lee and D. Sun. High-efficiency quantitative control of mitochondrial transfer based on droplet microfluidics and its application on muscle regeneration. Science Advances 2022, 8, 9245
[4] J. Suh, N.K. Kim, W. Shim, S.H. Lee, H.J. Kim, E. Moon, H. Sesaki, J.H. Jang, J.E. Kim and Y.S. Lee. Cell Metabolism 2023, 35 345-360
[5] D.M. Copolovici, K. Langel, E. Eriste, and U. Langel. Cell-Penetrating Peptides: Design, Synthesis, and Applications. ACS Nano2014, 8 (3), 1972-1944